


Summary
The Triassic (/traɪˈæsɪk/ try-ASS-ik; sometimes symbolized 🝈)[8] is a geologic period and system which spans 50.5 million years from the end of the Permian Period 251.902 million years ago (Mya), to the beginning of the Jurassic Period 201.4 Mya.[9] The Triassic is the first and shortest period of the Mesozoic Era. Both the start and end of the period are marked by major extinction events.[10] The Triassic Period is subdivided into three epochs: Early Triassic, Middle Triassic and Late Triassic.
| Triassic | |||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 A map of the world as it appeared during the Late Triassic, c. 220 Ma | |||||||||||||||||||||||
| Chronology | |||||||||||||||||||||||
| |||||||||||||||||||||||
| Etymology | |||||||||||||||||||||||
| Name formality | Formal | ||||||||||||||||||||||
| Usage information | |||||||||||||||||||||||
| Celestial body | Earth | ||||||||||||||||||||||
| Regional usage | Global (ICS) | ||||||||||||||||||||||
| Time scale(s) used | ICS Time Scale | ||||||||||||||||||||||
| Definition | |||||||||||||||||||||||
| Chronological unit | Period | ||||||||||||||||||||||
| Stratigraphic unit | System | ||||||||||||||||||||||
| Time span formality | Formal | ||||||||||||||||||||||
| Lower boundary definition | First appearance of the conodont Hindeodus parvus | ||||||||||||||||||||||
| Lower boundary GSSP | Meishan, Zhejiang, China 31°04′47″N 119°42′21″E / 31.0798°N 119.7058°E | ||||||||||||||||||||||
| Lower GSSP ratified | 2001[6] | ||||||||||||||||||||||
| Upper boundary definition | First appearance of the ammonite Psiloceras spelae tirolicum | ||||||||||||||||||||||
| Upper boundary GSSP | Kuhjoch section, Karwendel mountains, Northern Calcareous Alps, Austria 47°29′02″N 11°31′50″E / 47.4839°N 11.5306°E | ||||||||||||||||||||||
| Upper GSSP ratified | 2010[7] | ||||||||||||||||||||||
The Triassic began in the wake of the Permian–Triassic extinction event, which left the Earth's biosphere impoverished; it was well into the middle of the Triassic before life recovered its former diversity. Three categories of organisms can be distinguished in the Triassic record: survivors from the extinction event, new groups that flourished briefly, and other new groups that went on to dominate the Mesozoic Era. Reptiles, especially archosaurs, were the chief terrestrial vertebrates during this time. A specialized subgroup of archosaurs, called dinosaurs, first appeared in the Late Triassic but did not become dominant until the succeeding Jurassic Period.[11] Archosaurs that became dominant in this period were primarily pseudosuchians, relatives and ancestors of modern crocodilians, while some archosaurs specialized in flight, the first time among vertebrates, becoming the pterosaurs.
Therapsids, the dominant vertebrates of the preceding Permian period, declined throughout the period. The first true mammals, themselves a specialized subgroup of therapsids, also evolved during this period. The vast supercontinent of Pangaea dominated the globe during the Triassic, but in the following Jurassic period it began to gradually rift into two separate landmasses, Laurasia to the north and Gondwana to the south.
The global climate during the Triassic was mostly hot and dry,[12] with deserts spanning much of Pangaea's interior. However, the climate shifted and became more humid as Pangaea began to drift apart. The end of the period was marked by yet another major mass extinction, the Triassic–Jurassic extinction event, that wiped out many groups, including most pseudosuchians, and allowed dinosaurs to assume dominance in the Jurassic.
Etymology edit
The Triassic was named in 1834 by Friedrich August von Alberti, after a succession of three distinct rock layers (Greek triás meaning 'triad') that are widespread in southern Germany: the lower Buntsandstein (colourful sandstone), the middle Muschelkalk (shell-bearing limestone) and the upper Keuper (coloured clay).[13]
-
 Early Triassic sandstone (Buntsandstein) near Stadtroda, Germany
Early Triassic sandstone (Buntsandstein) near Stadtroda, Germany -
 Middle Triassic Muschelkalk (shell-bearing limestone) near Dörzbach, Germany
Middle Triassic Muschelkalk (shell-bearing limestone) near Dörzbach, Germany -




Dating and subdivisions edit
On the geologic time scale, the Triassic is usually divided into Early, Middle, and Late Triassic Epochs, and the corresponding rocks are referred to as Lower, Middle, or Upper Triassic. The faunal stages from the youngest to oldest are:
| Series/Epoch | Faunal stage | Time span |
|---|---|---|
| Upper/Late Triassic (Tr3) | Rhaetian | (208.5 – 201.4 ± 0.2 Mya) |
| Norian | (227 – 208.5 Mya) | |
| Carnian | (237 – 227 Mya) | |
| Middle Triassic (Tr2) | Ladinian | (242 – 237 Mya) |
| Anisian | (247.2 – 242 Mya) | |
| Lower/Early Triassic (Scythian) | Olenekian | (251.2 – 247.2 Mya) |
| Induan | (251.902 ± 0.024 – 251.2 Mya) |
Paleogeography edit

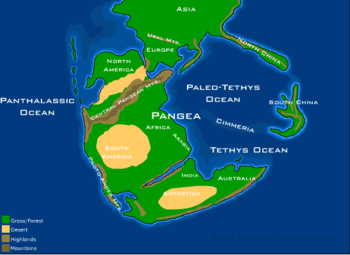
During the Triassic, almost all the Earth's land mass was concentrated into a single supercontinent, Pangaea (lit. 'entire land').[14] This supercontinent was more-or-less centered on the equator and extended between the poles, though it did drift northwards as the period progressed. Southern Pangea, also known as Gondwana, was made up by closely-appressed cratons corresponding to modern South America, Africa, Madagascar, India, Antarctica, and Australia. North Pangea, also known as Laurussia or Laurasia, corresponds to modern-day North America and the fragmented predecessors of Eurasia.
The western edge of Pangea lay at the margin of an enormous ocean, Panthalassa (lit. 'entire sea'), which roughly corresponds to the modern Pacific Ocean. Practically all deep-ocean crust present during the Triassic has been recycled through the subduction of oceanic plates, so very little is known about the open ocean from this time period. Most information on Panthalassan geology and marine life is derived from island arcs and rare seafloor sediments accreted onto surrounding land masses, such as present-day Japan and western North America.
The eastern edge of Pangea was encroached upon by a pair of extensive oceanic basins: The Neo-Tethys (or simply Tethys) and Paleo-Tethys Oceans. These extended from China to Iberia, hosting abundant marine life along their shallow tropical peripheries. They were divided from each other by a long string of microcontinents known as the Cimmerian terranes. Cimmerian crust had detached from Gondwana in the early Permian and drifted northwards during the Triassic, enlarging the Neo-Tethys Ocean which formed in their wake. At the same time, they forced the Paleo-Tethys Ocean to shrink as it was being subducted under Asia. By the end of the Triassic, the Paleo-Tethys Ocean occupied a small area and the Cimmerian terranes began to collide with southern Asia. This collision, known as the Cimmerian Orogeny, continued into the Jurassic and Cretaceous to produce a chain of mountain ranges stretching from Turkey to Malaysia.[15][16]

Pangaea was fractured by widespread faulting and rift basins during the Triassic—especially late in that period—but had not yet separated. The first nonmarine sediments in the rift that marks the initial break-up of Pangaea, which separated eastern North America from Morocco, are of Late Triassic age; in the United States, these thick sediments comprise the Newark Supergroup.[18] Rift basins are also common in South America, Europe, and Africa. Terrestrial environments are particularly well-represented in the South Africa,[19] Russia, central Europe, and the southwest United States. Terrestrial Triassic biostratigraphy is mostly based on terrestrial and freshwater tetrapods, as well as conchostracans ("clam shrimps"), a type of fast-breeding crustacean which lived in lakes and hypersaline environments.
Because a supercontinent has less shoreline compared to a series of smaller continents, Triassic marine deposits are relatively uncommon on a global scale. A major exception is in Western Europe, where the Triassic was first studied. The northeastern margin of Gondwana was a stable passive margin along the Neo-Tethys Ocean, and marine sediments have been preserved in parts of northern India and Arabia.[16] In North America, marine deposits are limited to a few exposures in the west.
edit
During the Triassic peneplains are thought to have formed in what is now Norway and southern Sweden.[20][21][22] Remnants of this peneplain can be traced as a tilted summit accordance in the Swedish West Coast.[20] In northern Norway Triassic peneplains may have been buried in sediments to be then re-exposed as coastal plains called strandflats.[21] Dating of illite clay from a strandflat of Bømlo, southern Norway, have shown that landscape there became weathered in Late Triassic times (c. 210 million years ago) with the landscape likely also being shaped during that time.[23]
Paleooceanography edit
Eustatic sea level in the Triassic was consistently low compared to the other geological periods. The beginning of the Triassic was around present sea level, rising to about 10–20 metres (33–66 ft) above present-day sea level during the Early and Middle Triassic. Sea level rise accelerated in the Ladinian, culminating with a sea level up to 50 metres (164 ft) above present-day levels during the Carnian. Sea level began to decline in the Norian, reaching a low of 50 metres (164 ft) below present sea level during the mid-Rhaetian. Low global sea levels persisted into the earliest Jurassic. The long-term sea level trend is superimposed by 22 sea level drop events widespread in the geologic record, mostly of minor (less than 25-metre (82 ft)) and medium (25–75-metre (82–246 ft)) magnitudes. A lack of evidence for Triassic continental ice sheets suggest that glacial eustasy is unlikely to be the cause of these changes.[24]
Climate edit
The Triassic continental interior climate was generally hot and dry, so that typical deposits are red bed sandstones and evaporites. There is no evidence of glaciation at or near either pole; in fact, the polar regions were apparently moist and temperate, providing a climate suitable for forests and vertebrates, including reptiles. Pangaea's large size limited the moderating effect of the global ocean; its continental climate was highly seasonal, with very hot summers and cold winters.[25] The strong contrast between the Pangea supercontinent and the global ocean triggered intense cross-equatorial monsoons,[25] sometimes referred to as the Pangean megamonsoons.[26]
The Triassic may have mostly been a dry period, but evidence exists that it was punctuated by several episodes of increased rainfall in tropical and subtropical latitudes of the Tethys Sea and its surrounding land.[27] Sediments and fossils suggestive of a more humid climate are known from the Anisian to Ladinian of the Tethysian domain, and from the Carnian and Rhaetian of a larger area that includes also the Boreal domain (e.g., Svalbard Islands), the North American continent, the South China block and Argentina. The best-studied of such episodes of humid climate, and probably the most intense and widespread, was the Carnian Pluvial Event.
Early Triassic edit
The Early Triassic was the hottest portion of the entire Phanerozoic, seeing as it occurred during and immediately after the discharge of titanic volumes of greenhouse gases from the Siberian Traps. The Early Triassic began with the Permian-Triassic Thermal Maximum (PTTM) and was followed by the brief Dienerian Cooling (DC) from 251 to 249 Ma, which was in turn followed by the Latest Smithian Thermal Maximum (LSTT) around 249 to 248 Ma. During the Latest Olenekian Cooling (LOC), from 248 to 247 Ma, temperatures cooled by about 6 °C.[28]
Middle Triassic edit
The Middle Triassic was cooler than the Early Triassic, with temperatures falling over most of the Anisian, with the exception of a warming spike in the latter portion of the stage.[29] From 242 to 233 Ma, the Ladinian-Carnian Cooling (LCC) ensued.[28]
Late Triassic edit
At the beginning of the Carnian, global temperatures continued to be relatively cool.[30] The eruption of the Wrangellia Large Igneous Province around 234 Ma caused abrupt global warming, terminating the cooling trend of the LCC.[31] This warming was responsible for the Carnian Pluvial Event and resulted in an episode of widespread global humidity.[32] The CPE ushered in the Mid-Carnian Warm Interval (MCWI), which lasted from 234 to 227 Ma.[28] At the Carnian-Norian boundary occurred a positive δ13C excursion believed to signify an increase in organic carbon burial.[33] From 227 to 217 Ma, there was a relatively cool period known as the Early Norian Cool Interval (ENCI), after which occurred the Mid-Norian Warm Interval (MNWI) from 217 to 209 Ma. The MNWI was briefly interrupted around 214 Ma by a cooling possibly related to the Manicouagan impact.[28] Around 212 Ma, a 10 Myr eccentricity maximum caused a paludification of Pangaea and a reduction in the size of arid climatic zones.[34] The Rhaetian Cool Interval (RCI) lasted from 209 to 201 Ma.[28] At the terminus of the Triassic, there was an extreme warming event referred to as the End-Triassic Thermal Event (ETTE), which was responsible for the Triassic-Jurassic mass extinction.[28] Bubbles of carbon dioxide in basaltic rocks dating back to the end of the Triassic indicate that volcanic activity from the Central Atlantic Magmatic Province helped trigger climate change in the ETTE.[35]
Flora edit

Land plants edit
During the Early Triassic, lycophytes, particularly those of the order Isoetales (which contains living quillworts), rose to promience due to the environmental instability following the Permian-Triassic extinction, with one particularly notable example being the genus Pleuromeia, which grew in columnar like fashion, sometimes reaching a height of 2 metres (6.6 ft). The relevance of lycophytes declined from the Middle Triassic onwards, following the return of more stable environmental conditions.[36]
While having first appeared during the Permian, the extinct seed plant group Bennettitales first became a prominent element in global floras during the Late Triassic, a position they would hold for much of the Mesozoic.[37] In the Southern Hemisphere landmasses of Gondwana, the tree Dicroidium, an extinct "seed fern" belong to the order Corystospermales was a dominant element in forest habitats across the region during the Middle-Late Triassic.[38] During the Late Triassic, the Ginkgoales (which today are represented by only a single species, Ginkgo biloba) underwent considerable diversification.[39] Conifers were abundant during the Triassic, and included the Voltziales (which contains various lineages, probably including those ancestral to modern conifers),[40] as well as the extinct family Cheirolepidiaceae, which first appeared in the Late Triassic, and would be prominent throught most of the rest of the Mesozoic.[41]
Coal edit

No known coal deposits date from the start of the Triassic Period. This is known as the Early Triassic "coal gap" and can be seen as part of the Permian–Triassic extinction event.[43] Possible explanations for the coal gap include sharp drops in sea level at the time of the Permo-Triassic boundary;[44] acid rain from the Siberian Traps eruptions or from an impact event that overwhelmed acidic swamps; climate shift to a greenhouse climate that was too hot and dry for peat accumulation; evolution of fungi or herbivores that were more destructive of wetlands; the extinction of all plants adapted to peat swamps, with a hiatus of several million years before new plant species evolved that were adapted to peat swamps;[43] or soil anoxia as oxygen levels plummeted.[45]
Phytoplankton edit
Before the Permian extinction, Archaeplastida (red and green algae) had been the major marine phytoplanktons since about 659–645 million years ago,[46] when they replaced marine planktonic cyanobacteria, which first appeared about 800 million years ago, as the dominant phytoplankton in the oceans.[47] In the Triassic, secondary endosymbiotic algae became the most important plankton.[48]
Fauna edit

Marine invertebrates edit
In marine environments, new modern types of corals appeared in the Early Triassic, forming small patches of reefs of modest extent compared to the great reef systems of Devonian or modern times. At the end of the Carnian, a reef crisis occurred in South China.[49] Serpulids appeared in the Middle Triassic.[50] Microconchids were abundant. The shelled cephalopods called ammonites recovered, diversifying from a single line that survived the Permian extinction. Bivalves began to rapidly diversify during the Middle Triassic, becoming highly abundant in the oceans.[51]
Fish edit

In the wake of the Permian-Triassic mass extinction event, the fish fauna was remarkably uniform, with many families and genera exhibiting a cosmopolitan distribution.[52] Coelacanths show their highest post-Devonian diversity in the Early Triassic.[53] Ray-finned fishes (actinopterygians) went through a remarkable diversification in the beginning of the Triassic, leading to peak diversity during the Middle Triassic; however, the pattern of this diversification is still not well understood due to a taphonomic megabias.[54] The first stem-group teleosts appeared during the Triassic (teleosts are by far the most diverse group of fish today).[52] Predatory actinopterygians such as saurichthyids and birgeriids, some of which grew over 1.2 m (3.9 ft) in length, appeared in the Early Triassic and became widespread and successful during the period as a whole.[55] Lakes and rivers were populated by lungfish (Dipnoi), such as Ceratodus, which are mainly known from the dental plates, abundant in the fossils record.[56] Hybodonts, a group of shark-like cartilaginous fish, were dominant in both freshwater and marine environments throughout the Triassic.[57] Last survivors of the mainly Palaeozoic Eugeneodontida are known from the Early Triassic.[58]
Amphibians edit
Temnospondyl amphibians were among those groups that survived the Permian–Triassic extinction. Once abundant in both terrestrial and aquatic environments, the terrestrial species had mostly died out during the extinction event. The Triassic survivors were aquatic or semi-aquatic, and were represented by Tupilakosaurus, Thabanchuia, Branchiosauridae and Micropholis, all of which died out in Early Triassic, and the successful Stereospondyli, with survivors into the Cretaceous Period. The largest Triassic stereospondyls, such as Mastodonsaurus, were up to 4 to 6 metres (13 to 20 ft) in length.[59][60] Some lineages (e.g. trematosaurs) flourished briefly in the Early Triassic, while others (e.g. capitosaurs) remained successful throughout the whole period, or only came to prominence in the Late Triassic (e.g. Plagiosaurus, metoposaurs).
The first Lissamphibians (modern amphibians) appear in the Triassic, with the progenitors of the first frogs already present by the Early Triassic. However, the group as a whole did not become common until the Jurassic, when the temnospondyls had become very rare.
Most of the Reptiliomorpha, stem-amniotes that gave rise to the amniotes, disappeared in the Triassic, but two water-dwelling groups survived: Embolomeri that only survived into the early part of the period, and the Chroniosuchia, which survived until the end of the Triassic.
Reptiles edit
Archosauromorphs edit
The Permian–Triassic extinction devastated terrestrial life. Biodiversity rebounded as the surviving species repopulated empty terrain, but these were short-lived. Diverse communities with complex food-web structures took 30 million years to reestablish.[10][61] Archosauromorph reptiles, which had already appeared and diversified to an extent in the Permian Period, exploded in diversity as an adaptive radiation in response to the Permian-Triassic mass extinction. By the Early Triassic, several major archosauromorph groups had appeared. Long-necked, lizard-like early archosauromorphs were known as protorosaurs, which is likely a paraphyletic group rather than a true clade. Tanystropheids were a family of protorosaurs which elevated their neck size to extremes, with the largest genus Tanystropheus having a neck longer than its body. The protorosaur family Sharovipterygidae used their elongated hindlimbs for gliding. Other archosauromorphs, such as rhynchosaurs and allokotosaurs, were mostly stocky-bodied herbivores with specialized jaw structures.
Rhynchosaurs, barrel-gutted herbivores, thrived for only a short period of time, becoming extinct about 220 million years ago. They were exceptionally abundant in the middle of the Triassic, as the primary large herbivores in many Carnian-age ecosystems. They sheared plants with premaxillary beaks and plates along the upper jaw with multiple rows of teeth. Allokotosaurs were iguana-like reptiles, including Trilophosaurus (a common Late Triassic reptile with three-crowned teeth), Teraterpeton (which had a long beak-like snout), and Shringasaurus (a horned herbivore which reached a body length of 3–4 metres (9.8–13.1 ft).
One group of archosauromorphs, the archosauriforms, were distinguished by their active predatory lifestyle, with serrated teeth and upright limb postures. Archosauriforms were diverse in the Triassic, including various terrestrial and semiaquatic predators of all shapes and sizes. The large-headed and robust erythrosuchids were among the dominant carnivores in the early Triassic. Phytosaurs were a particularly common group which prospered during the Late Triassic. These long-snouted and semiaquatic predators resemble living crocodiles and probably had a similar lifestyle, hunting for fish and small reptiles around the water's edge. However, this resemblance is only superficial and is a prime-case of convergent evolution.
True archosaurs appeared in the early Triassic, splitting into two branches: Avemetatarsalia (the ancestors to birds) and Pseudosuchia (the ancestors to crocodilians). Avemetatarsalians were a minor component of their ecosystems, but eventually produced the earliest pterosaurs and dinosaurs in the Late Triassic. Early long-tailed pterosaurs appeared in the Norian and quickly spread worldwide. Triassic dinosaurs evolved in the Carnian and include early sauropodomorphs and theropods. Most Triassic dinosaurs were small predators and only a few were common, such as Coelophysis, which was 1 to 2 metres (3.3 to 6.6 ft) long. Triassic sauropodomorphs primarily inhabited cooler regions of the world.[62]
The large predator Smok was most likely also an archosaur, but it is uncertain if it was a primitive dinosaur or a pseudosuchian.
Pseudosuchians were far more ecologically dominant in the Triassic, including large herbivores (such as aetosaurs), large carnivores ("rauisuchians"), and the first crocodylomorphs ("sphenosuchians"). Aetosaurs were heavily-armored reptiles that were common during the last 30 million years of the Late Triassic until they died out at the Triassic-Jurassic extinction. Most aetosaurs were herbivorous and fed on low-growing plants, but some may have eaten meat. "rauisuchians" (formally known as paracrocodylomorphs) were the keystone predators of most Triassic terrestrial ecosystems. Over 25 species have been found, including giant quadrupedal hunters, sleek bipedal omnivores, and lumbering beasts with deep sails on their backs. They probably occupied the large-predator niche later filled by theropods. "Rauisuchians" were ancestral to small, lightly-built crocodylomorphs, the only pseudosuchians which survived into the Jurassic.
-
 Tanystropheus, a long-necked tanystropheid
Tanystropheus, a long-necked tanystropheid -
 Proterosuchus, a crocodile-like early archosauriform from the Early Triassic
Proterosuchus, a crocodile-like early archosauriform from the Early Triassic -
 Staurikosaurus, one of the earliest dinosaurs, a member of the Triassic family Herrerasauridae
Staurikosaurus, one of the earliest dinosaurs, a member of the Triassic family Herrerasauridae -

-
 Plateosaurus was one of the largest of early sauropodomorphs, or "prosauropods", of the Late Triassic
Plateosaurus was one of the largest of early sauropodomorphs, or "prosauropods", of the Late Triassic -
 Coelophysis was one of the most abundant theropod dinosaurs in the Late Triassic
Coelophysis was one of the most abundant theropod dinosaurs in the Late Triassic



Marine reptiles edit

There were many types of marine reptiles. These included the Sauropterygia, which featured pachypleurosaurus and nothosaurs (both common during the Middle Triassic, especially in the Tethys region), placodonts, the earliest known herbivorous marine reptile Atopodentatus, and the first plesiosaurs. The first of the lizardlike Thalattosauria (askeptosaurs) and the highly successful ichthyosaurs, which appeared in Early Triassic seas soon diversified, and some eventually developed to huge size during the Late Triassic.
Other reptiles edit
Among other reptiles, the earliest turtles, like Proganochelys and Proterochersis, appeared during the Norian Age (Stage) of the Late Triassic Period. The Lepidosauromorpha, specifically the Sphenodontia, are first found in the fossil record of the earlier Carnian Age, though the earliest lepidosauromorphs likely occurred in the Permian. The Procolophonidae, the last surviving parareptiles, were an important group of small lizard-like herbivores. The drepanosaurs were a clade of unusual, chameleon-like arboreal reptiles with birdlike heads and specialised claws.
Synapsids edit
Three therapsid groups survived into the Triassic: dicynodonts, therocephalians, and cynodonts. The cynodont Cynognathus was a characteristic top predator in the Olenekian and Anisian of Gondwana. Both kannemeyeriiform dicynodonts and gomphodont cynodonts remained important herbivores during much of the period. Therocephalians included both large predators (Moschorhinus) and herbivorous forms (bauriids) until their extinction midway through the period. Ecteniniid cynodonts played a role as large-sized, cursorial predators in the Late Triassic. During the Carnian (early part of the Late Triassic), some advanced cynodonts gave rise to the first mammals.
During the Triassic, archosaurs displaced therapsids as the largest and most ecologically prolific terrestrial amniotes. This "Triassic Takeover" may have contributed to the evolution of mammals by forcing the surviving therapsids and their mammaliaform successors to live as small, mainly nocturnal insectivores. Nocturnal life may have forced the mammaliaforms to develop fur and a higher metabolic rate.[64]
-
 Lystrosaurus was a widespread dicynodont and the most common land vertebrate during the Early Triassic, after animal life had been greatly diminished
Lystrosaurus was a widespread dicynodont and the most common land vertebrate during the Early Triassic, after animal life had been greatly diminished -



Lagerstätten edit
Two Early Triassic lagerstätten (high-quality fossil beds), the Dienerian aged Guiyang biota[65] and the earliest Spathian aged Paris biota[66] stand out due to their exceptional preservation and diversity. They represent the earliest lagerstätten of the Mesozoic era and provide insight into the biotic recovery from the Permian-Triassic mass extinction event.
The Monte San Giorgio lagerstätte, now in the Lake Lugano region of northern Italy and southern Switzerland, was in Middle Triassic times a lagoon behind reefs with an anoxic bottom layer, so there were no scavengers and little turbulence to disturb fossilization, a situation that can be compared to the better-known Jurassic Solnhofen Limestone lagerstätte. The remains of fish and various marine reptiles (including the common pachypleurosaur Neusticosaurus, and the bizarre long-necked archosauromorph Tanystropheus), along with some terrestrial forms like Ticinosuchus and Macrocnemus, have been recovered from this locality. All these fossils date from the Anisian and Ladinian ages (about 242 Ma ago).
Triassic–Jurassic extinction event edit

The Triassic Period ended with a mass extinction, which was particularly severe in the oceans; the conodonts disappeared, as did all the marine reptiles except ichthyosaurs and plesiosaurs. Invertebrates like brachiopods and molluscs (such as gastropods) were severely affected. In the oceans, 22% of marine families and possibly about half of marine genera went missing.
Though the end-Triassic extinction event was not equally devastating in all terrestrial ecosystems, several important clades of crurotarsans (large archosaurian reptiles previously grouped together as the thecodonts) disappeared, as did most of the large labyrinthodont amphibians, groups of small reptiles, and most synapsids. Some of the early, primitive dinosaurs also became extinct, but more adaptive ones survived to evolve into the Jurassic. Surviving plants that went on to dominate the Mesozoic world included modern conifers and cycadeoids.
The cause of the Late Triassic extinction is uncertain. It was accompanied by huge volcanic eruptions that occurred as the supercontinent Pangaea began to break apart about 202 to 191 million years ago (40Ar/39Ar dates),[67] forming the Central Atlantic Magmatic Province (CAMP),[68] one of the largest known inland volcanic events since the planet had first cooled and stabilized. Other possible but less likely causes for the extinction events include global cooling or even a bolide impact, for which an impact crater containing Manicouagan Reservoir in Quebec, Canada, has been singled out. However, the Manicouagan impact melt has been dated to 214±1 Mya. The date of the Triassic-Jurassic boundary has also been more accurately fixed recently, at 201.4 Mya. Both dates are gaining accuracy by using more accurate forms of radiometric dating, in particular the decay of uranium to lead in zircons formed at time of the impact. So, the evidence suggests the Manicouagan impact preceded the end of the Triassic by approximately 10±2 Ma. It could not therefore be the immediate cause of the observed mass extinction.[69]

The number of Late Triassic extinctions is disputed. Some studies suggest that there are at least two periods of extinction towards the end of the Triassic, separated by 12 to 17 million years. But arguing against this is a recent study of North American faunas. In the Petrified Forest of northeast Arizona there is a unique sequence of late Carnian-early Norian terrestrial sediments. An analysis in 2002 found no significant change in the paleoenvironment.[70] Phytosaurs, the most common fossils there, experienced a change-over only at the genus level, and the number of species remained the same. Some aetosaurs, the next most common tetrapods, and early dinosaurs, passed through unchanged. However, both phytosaurs and aetosaurs were among the groups of archosaur reptiles completely wiped out by the end-Triassic extinction event.
It seems likely then that there was some sort of end-Carnian extinction, when several herbivorous archosauromorph groups died out, while the large herbivorous therapsids—the kannemeyeriid dicynodonts and the traversodont cynodonts—were much reduced in the northern half of Pangaea (Laurasia).
These extinctions within the Triassic and at its end allowed the dinosaurs to expand into many niches that had become unoccupied. Dinosaurs became increasingly dominant, abundant and diverse, and remained that way for the next 150 million years. The true "Age of Dinosaurs" is during the following Jurassic and Cretaceous periods, rather than the Triassic.
See also edit
- Geologic time scale
- List of fossil sites (with link directory)
- Triassic land vertebrate faunachrons
- Phylloceratina
- Dinosaurs
Notes edit
- ^ Widmann, Philipp; Bucher, Hugo; Leu, Marc; et al. (2020). "Dynamics of the Largest Carbon Isotope Excursion During the Early Triassic Biotic Recovery". Frontiers in Earth Science. 8 (196): 196. Bibcode:2020FrEaS...8..196W. doi:10.3389/feart.2020.00196.
- ^ McElwain, J. C.; Punyasena, S. W. (2007). "Mass extinction events and the plant fossil record". Trends in Ecology & Evolution. 22 (10): 548–557. doi:10.1016/j.tree.2007.09.003. PMID 17919771.
- ^ Retallack, G. J.; Veevers, J.; Morante, R. (1996). "Global coal gap between Permian–Triassic extinctions and middle Triassic recovery of peat forming plants". GSA Bulletin. 108 (2): 195–207. Bibcode:1996GSAB..108..195R. doi:10.1130/0016-7606(1996)108<0195:GCGBPT>2.3.CO;2. Retrieved 2007-09-29.
- ^ Payne, J. L.; Lehrmann, D. J.; Wei, J.; Orchard, M. J.; Schrag, D. P.; Knoll, A. H. (2004). "Large Perturbations of the Carbon Cycle During Recovery from the End-Permian Extinction". Science. 305 (5683): 506–9. Bibcode:2004Sci...305..506P. doi:10.1126/science.1097023. PMID 15273391. S2CID 35498132.
- ^ Ogg, James G.; Ogg, Gabi M.; Gradstein, Felix M. (2016). "Triassic". A Concise Geologic Time Scale: 2016. Elsevier. pp. 133–149. ISBN 978-0-444-63771-0.
- ^ Hongfu, Yin; Kexin, Zhang; Jinnan, Tong; Zunyi, Yang; Shunbao, Wu (June 2001). "The Global Stratotype Section and Point (GSSP) of the Permian-Triassic Boundary" (PDF). Episodes. 24 (2): 102–114. doi:10.18814/epiiugs/2001/v24i2/004. Retrieved 8 December 2020.
- ^ Hillebrandt, A.v.; Krystyn, L.; Kürschner, W. M.; et al. (September 2013). "The Global Stratotype Sections and Point (GSSP) for the base of the Jurassic System at Kuhjoch (Karwendel Mountains, Northern Calcareous Alps, Tyrol, Austria)". Episodes. 36 (3): 162–198. CiteSeerX 10.1.1.736.9905. doi:10.18814/epiiugs/2013/v36i3/001. S2CID 128552062.
- ^ "Triassic". Dictionary.com Unabridged (Online). n.d.
- ^ "International Stratigraphic Chart" (PDF). International Commission on Stratigraphy. June 2023. Retrieved 19 July 2023.
- ^ a b Sahney, S. & Benton, M.J. (2008). "Recovery from the most profound mass extinction of all time". Proceedings of the Royal Society B: Biological Sciences. 275 (1636): 759–765. doi:10.1098/rspb.2007.1370. PMC 2596898. PMID 18198148.
- ^ Brusatte, S. L.; Benton, M. J.; Ruta, M.; Lloyd, G. T. (2008-09-12). "Superiority, Competition, and Opportunism in the Evolutionary Radiation of Dinosaurs" (PDF). Science. 321 (5895): 1485–1488. Bibcode:2008Sci...321.1485B. doi:10.1126/science.1161833. hdl:20.500.11820/00556baf-6575-44d9-af39-bdd0b072ad2b. PMID 18787166. S2CID 13393888. Archived from the original (PDF) on 2014-06-24. Retrieved 2012-01-14.
- ^ "'Lethally Hot' Earth Was Devoid of Life – Could It Happen Again?". nationalgeographic.com. 19 October 2012. Archived from the original on October 20, 2012.
- ^ Friedrich von Alberti, Beitrag zu einer Monographie des bunten Sandsteins, Muschelkalks und Keupers, und die Verbindung dieser Gebilde zu einer Formation [Contribution to a monograph on the colored sandstone, shell limestone and mudstone, and the joining of these structures into one formation] (Stuttgart and Tübingen, (Germany): J. G. Cotta, 1834). Alberti coined the term "Trias" on page 324 :
"… bunter Sandstein, Muschelkalk und Keuper das Resultat einer Periode, ihre Versteinerungen, um mich der Worte E. de Beaumont's zu bedeinen, die Thermometer einer geologischen Epoche seyen, … also die bis jezt beobachtete Trennung dieser Gebilde in 3 Formationen nicht angemessen, und es mehr dem Begriffe Formation entsprechend sey, sie zu einer Formation, welche ich vorläufig Trias nennen will, zu verbinden."
( … colored sandstone, shell limestone, and mudstone are the result of a period; their fossils are, to avail myself of the words of E. de Beaumont, the thermometer of a geologic epoch; … thus the separation of these structures into 3 formations, which has been maintained until now, isn't appropriate, and it is more consistent with the concept of "formation" to join them into one formation, which for now I will name "trias".) - ^ Forte, Giuseppa; Kustatscher, Evelyn; Preto, Nereo (1 September 2022). "Carbon (δ13C) isotope variations indicate climate shifts and reflect plant habitats in the Middle Triassic (Anisian, Pelsonian) succession at Kühwiesenkopf / Monte Prà della Vacca (Dolomites, Northeast Italy)". Palaeogeography, Palaeoclimatology, Palaeoecology. 601: 111098. Bibcode:2022PPP...60111098F. doi:10.1016/j.palaeo.2022.111098. S2CID 249483335. Retrieved 1 December 2022.
- ^ Mazaheri-Johari, Mina; Roghi, Guido; Caggiati, Marcello; Kustatscher, Evelyn; Ghasemi-Nejad, Ebrahim; Zanchi, Andrea; Gianolla, Piero (15 January 2022). "Disentangling climate signal from tectonic forcing: The Triassic Aghdarband Basin (Turan Domain, Iran)". Palaeogeography, Palaeoclimatology, Palaeoecology. 586: 110777. Bibcode:2022PPP...58610777M. doi:10.1016/j.palaeo.2021.110777. S2CID 244696034. Retrieved 10 January 2023.
- ^ a b Cocks, L. Robin M. & Torsvik, Trond H., eds. (2016). "Triassic". Earth History and Palaeogeography. Cambridge, UK: Cambridge University Press. pp. 195–207. doi:10.1017/9781316225523.012. ISBN 978-1-316-22552-3. Retrieved 2022-05-16.
- ^ Herbert, Chris; Helby, Robin (1980). A Guide to the Sydney basin. Maitland, NSW: Geological Survey of NSW. p. 582. ISBN 978-0-7240-1250-3.
- ^ "Lecture 10 – Triassic: Newark, Chinle". rainbow.ldeo.columbia.edu.
- ^ Jacobs, Louis L. (1997). "African Dinosaurs". In Currie, Phillip J.; Padian, Kevin (eds.). Encyclopedia of Dinosaurs. Academic Press. pp. 2–4.
- ^ a b Lidmar-Bergström, Karna (1993). "Denudation surfaces and tectonics in the southernmost part of the Baltic Shield". Precambrian Research. 64 (1–4): 337–345. Bibcode:1993PreR...64..337L. doi:10.1016/0301-9268(93)90086-h.
- ^ a b Olesen, Odleiv; Kierulf, Halfdan Pascal; Brönner, Marco; Dalsegg, Einar; Fredin, Ola; Solbakk, Terje (2013). "Deep weathering, neotectonics and strandflat formation in Nordland, northern Norway". Norwegian Journal of Geology. 93: 189–213.
- ^ Japsen, Peter; Green, Paul F; Bonow, Johan M; Erlström, Mikael (2016). "Episodic burial and exhumation of the southern Baltic Shield: Epeirogenic uplifts during and after break-up of Pangaea". Gondwana Research. 35: 357–77. Bibcode:2016GondR..35..357J. doi:10.1016/j.gr.2015.06.005.
- ^ Fredin, Ola; Viola, Giulio; Zwingmann, Horst; Sørlie, Ronald; Brönner, Marco; Lie, Jan-Erik; et al. (2017). "The inheritance of a Mesozoic landscape in western Scandinavia". Nature. 8: 14879. Bibcode:2017NatCo...814879F. doi:10.1038/ncomms14879. PMC 5477494. PMID 28452366.
- ^ Haq, Bilal U. (December 2018). "Triassic eustatic variations reexamined". GSA Today. 28 (12). Geological Society of America: 4–9. Bibcode:2018GSAT...28l...4H. doi:10.1130/GSATG381A.1. S2CID 134477691.
- ^ a b Stanley, 452–53.
- ^ Zeng, Zhiwei; Zhu, Hongtao; Yang, Xianghua; Zeng, Hongliu; Hu, Xiaolin; Xia, Chenchen (May 2019). "The Pangaea Megamonsoon records: Evidence from the Triassic Mungaroo Formation, Northwest Shelf of Australia". Gondwana Research. 69: 1–24. Bibcode:2019GondR..69....1Z. doi:10.1016/j.gr.2018.11.015. S2CID 134145664. Retrieved 9 January 2023.
- ^ Preto, N.; Kustatscher, E.; Wignall, P. B. (2010). "Triassic climates – State of the art and perspectives". Palaeogeography, Palaeoclimatology, Palaeoecology. 290 (1–4): 1–10. Bibcode:2010PPP...290....1P. doi:10.1016/j.palaeo.2010.03.015.
- ^ a b c d e f Scotese, Christopher Robert; Song, Haijun; Mills, Benjamin J. W.; van der Meer, Douwe G. (1 April 2021). "Phanerozoic paleotemperatures: The earth's changing climate during the last 540 million years". Earth-Science Reviews. 215: 103503. Bibcode:2021ESRv..21503503S. doi:10.1016/j.earscirev.2021.103503. ISSN 0012-8252. S2CID 233579194. Retrieved 22 September 2023.
- ^ Trotter, Julie A.; Williams, Ian S.; Nicora, Alda; Mazza, Michele; Rigo, Manuel (April 2015). "Long-term cycles of Triassic climate change: a new δ18O record from conodont apatite". Earth and Planetary Science Letters. 415: 165–174. Bibcode:2015E&PSL.415..165T. doi:10.1016/j.epsl.2015.01.038. Retrieved 22 September 2023.
- ^ Dal Corso, Jacopo; Mills, Benjamin J.W.; Chu, Daoliang; Newton, Robert J.; Song, Haijun (15 January 2022). "Background Earth system state amplified Carnian (Late Triassic) environmental changes". Earth and Planetary Science Letters. 578: 117321. Bibcode:2022E&PSL.57817321D. doi:10.1016/j.epsl.2021.117321. S2CID 244847207. Retrieved 22 September 2023.
- ^ Dal Corso, J.; Mietto, P.; Newton, R. J.; Pancost, R. D.; Preto, N.; Roghi, G.; Wignall, P. B. (1 January 2012). "Discovery of a major negative 13C spike in the Carnian (Late Triassic) linked to the eruption of Wrangellia flood basalts". Geology. 40 (1): 79–82. Bibcode:2012Geo....40...79D. doi:10.1130/G32473.1. ISSN 0091-7613. Retrieved 22 September 2023.
- ^ Li, Liqin; Kürschner, Wolfram M.; Lu, Ning; Chen, Hongyu; An, Pengcheng; Wang, Yongdong (September 2022). "Palynological record of the Carnian Pluvial Episode from the northwestern Sichuan Basin, SW China". Review of Palaeobotany and Palynology. 304: 104704. Bibcode:2022RPaPa.30404704L. doi:10.1016/j.revpalbo.2022.104704. S2CID 249528886. Retrieved 22 September 2023.
- ^ Muttoni, Giovanni; Mazza, Michele; Mosher, David; Katz, Miriam E.; Kent, Dennis V.; Balini, Marco (1 April 2014). "A Middle–Late Triassic (Ladinian–Rhaetian) carbon and oxygen isotope record from the Tethyan Ocean". Palaeogeography, Palaeoclimatology, Palaeoecology. 399: 246–259. Bibcode:2014PPP...399..246M. doi:10.1016/j.palaeo.2014.01.018. ISSN 0031-0182. Retrieved 24 November 2023.
- ^ Ikeda, Masayuki; Ozaki, Kazumi; Legrand, Julien (23 July 2020). "Impact of 10-Myr scale monsoon dynamics on Mesozoic climate and ecosystems". Scientific Reports. 10 (1): 11984. Bibcode:2020NatSR..1011984I. doi:10.1038/s41598-020-68542-w. ISSN 2045-2322. PMC 7378230. PMID 32704030.
- ^ Capriolo, Manfredo; Marzoli, Andrea; Aradi, László E.; Callegaro, Sara; Dal Corso, Jacopo; Newton, Robert J.; Mills, Benjamin J. W.; Wignall, Paul B.; Bartoli, Omar; Baker, Don R.; Youbi, Nasrrddine; Remusat, Laurent; Spiess, Richard; Szabó, Csaba (7 April 2020). "Deep CO2 in the end-Triassic Central Atlantic Magmatic Province". Nature Communications. 11 (1): 1670. Bibcode:2020NatCo..11.1670C. doi:10.1038/s41467-020-15325-6. ISSN 2041-1723. PMC 7138847. PMID 32265448.
- ^ Looy, Cindy V.; van Konijnenburg-van Cittert, Johanna H. A.; Duijnstee, Ivo A. P. (2021-03-02). "Proliferation of Isoëtalean Lycophytes During the Permo-Triassic Biotic Crises: A Proxy for the State of the Terrestrial Biosphere". Frontiers in Earth Science. 9: 55. Bibcode:2021FrEaS...9...55L. doi:10.3389/feart.2021.615370. ISSN 2296-6463.
- ^ Blomenkemper, Patrick; Bäumer, Robert; Backer, Malte; Abu Hamad, Abdalla; Wang, Jun; Kerp, Hans; Bomfleur, Benjamin (2021). "Bennettitalean Leaves From the Permian of Equatorial Pangea—The Early Radiation of an Iconic Mesozoic Gymnosperm Group". Frontiers in Earth Science. 9: 162. Bibcode:2021FrEaS...9..162B. doi:10.3389/feart.2021.652699.
- ^ Mays, Chris; McLoughlin, Stephen (2020-02-25). "Caught between two mass extinctions: The rise and fall of Dicroidium". Deposits Mag. Retrieved 2023-09-23.
- ^ Zhou, Zhi-Yan (March 2009). "An overview of fossil Ginkgoales". Palaeoworld. 18 (1): 1–22. doi:10.1016/j.palwor.2009.01.001.
- ^ Forte, Giuseppa; Kustatscher, Evelyn; Van Konijnenburg-van Cittert, Johanna H. A. (2021-07-01). "Conifer Diversity in the Middle Triassic: New Data from the Fossillagerstätte Kühwiesenkopf/Monte Prà della Vacca (Pelsonian, Anisian) in the Dolomites (Northeastern Italy)". International Journal of Plant Sciences. 182 (6): 445–467. doi:10.1086/714280. ISSN 1058-5893.
- ^ Escapa, Ignacio; Leslie, Andrew (February 2017). "A new Cheirolepidiaceae (Coniferales) from the Early Jurassic of Patagonia (Argentina): Reconciling the records of impression and permineralized fossils". American Journal of Botany. 104 (2): 322–334. doi:10.3732/ajb.1600321. ISSN 0002-9122. PMID 28213347.
- ^ Hosher, WT Magaritz M Clark D (1987). "Events near the time of the Permian-Triassic boundary". Mod. Geol. 11: 155–80 [173–74].
- ^ a b Retallack, G. J.; Veevers, J. J.; Morante, R. (1996). "Global coal gap between Permian-Triassic extinction and Middle Triassic recovery of peat-forming plants". Bulletin of the Geological Society of America. 108 (2): 195–207. Bibcode:1996GSAB..108..195R. doi:10.1130/0016-7606(1996)108<0195:GCGBPT>2.3.CO;2.
- ^ Holser, WT; Schoenlaub, H-P; Klein, P; Attrep, M; Boeckelmann, Klaus; et al. (1989). "A unique geochemical record at the Permian/Triassic boundary". Nature. 337 (6202): 39 [42]. Bibcode:1989Natur.337...39H. doi:10.1038/337039a0. S2CID 8035040. Retrieved 24 November 2023.
- ^ Retallack, G.J.; Krull, E.S. (2006). "Carbon isotopic evidence for terminal-Permian methane outbursts and their role in extinctions of animals, plants, coral reefs, and peat swamps" (PDF). Geological Society of America Special Paper. 399: 249. doi:10.1130/2006.2399(12). ISBN 978-0-8137-2399-0. Retrieved 14 December 2020.
- ^ "How snowball Earth gave rise to complex life – Cosmos Magazine". 16 August 2017.
- ^ "December: Phytoplankton | News | University of Bristol".
- ^ "The rise of algae in Cryogenian oceans and the emergence of animals – ResearchGate".
- ^ Jin, Xin; Shi, Zhiqiang; Rigo, Manuel; Franceschi, Marco; Preto, Nereo (15 September 2018). "Carbonate platform crisis in the Carnian (Late Triassic) of Hanwang (Sichuan Basin, South China): Insights from conodonts and stable isotope data". Journal of Asian Earth Sciences. 164: 104–124. Bibcode:2018JAESc.164..104J. doi:10.1016/j.jseaes.2018.06.021. S2CID 134733944. Retrieved 22 September 2023.
- ^ Vinn, O.; Mutvei, H. (2009). "Calcareous tubeworms of the Phanerozoic" (PDF). Estonian Journal of Earth Sciences. 58 (4): 286–96. doi:10.3176/earth.2009.4.07. Retrieved 2012-09-16.
- ^ Komatsu, Toshifumi; Chen, Jin-hua; Cao, Mei-zhen; Stiller, Frank; Naruse, Hajime (10 June 2004). "Middle Triassic (Anisian) diversified bivalves: depositional environments and bivalve assemblages in the Leidapo Member of the Qingyan Formation, southern China". Palaeogeography, Palaeoclimatology, Palaeoecology. 208 (3–4): 227–223. Bibcode:2004PPP...208..207K. doi:10.1016/j.palaeo.2004.03.005. Retrieved 31 March 2023.
- ^ a b Romano, Carlo; Koot, Martha B.; Kogan, Ilja; Brayard, Arnaud; Minikh, Alla V.; Brinkmann, Winand; Bucher, Hugo; Kriwet, Jürgen (February 2016). "Permian-Triassic Osteichthyes (bony fishes): diversity dynamics and body size evolution". Biological Reviews. 91 (1): 106–47. doi:10.1111/brv.12161. PMID 25431138. S2CID 5332637.
- ^ Cavin, Lionel; Furrer, Heinz; Obrist, Christian (2013). "New coelacanth material from the Middle Triassic of eastern Switzerland, and comments on the taxic diversity of actinistans". Swiss Journal of Geoscience. 106 (2): 161–177. doi:10.1007/s00015-013-0143-7.
- ^ Romano, Carlo (January 2021). "A hiatus obscures the early evolution of Modern lineages of bony fishes". Frontiers in Earth Science. 8: 618853. doi:10.3389/feart.2020.618853.
- ^ Romano, Carlo; Jenks, James F.; Jattiot, Romain; Scheyer, Torsten M. (2017). "Marine Early Triassic Actinopterygii from Elko County (Nevada, USA): implications for the Smithian equatorial vertebrate eclipse". Journal of Paleontology. 91 (5): 1–22. Bibcode:2017JPal...91.1025R. doi:10.1017/jpa.2017.36.
- ^ Agnolin, F. L., Mateus O., Milàn J., Marzola M., Wings O., Adolfssen J. S., & Clemmensen L. B. (2018). Ceratodus tunuensis, sp. nov., a new lungfish (Sarcopterygii, Dipnoi) from the Upper Triassic of central East Greenland. Journal of Vertebrate PaleontologyJournal of Vertebrate Paleontology. e1439834
- ^ Kumar, Krishna; Bajpai, Sunil; Pandey, Pragya; Ghosh, Triparna; Bhattacharya, Debasish (2021-08-04). "Hybodont sharks from the Jurassic of Jaisalmer, western India". Historical Biology. 34 (6): 953–963. doi:10.1080/08912963.2021.1954920. ISSN 0891-2963. S2CID 238781606.
- ^ Mutter, Raoul J.; Neuman, Andrew G. (2008). "New eugeneodontid sharks from the Lower Triassic Sulphur Mountain Formation of Western Canada". In Cavin, L.; Longbottom, A.; Richter, M. (eds.). Fishes and the Break-up of Pangaea. Geological Society of London, Special Publications. Vol. 295. London: Geological Society of London. pp. 9–41. doi:10.1144/sp295.3. S2CID 130268582.
- ^ Wells, Kentwood D. (2010). The Ecology and Behavior of Amphibians. University of Chicago Press. ISBN 978-0-226-89333-4 – via Google Books.
- ^ Benton, Michael (2009). Vertebrate Palaeontology. John Wiley & Sons. ISBN 978-1-4051-4449-0 – via Google Books.
- ^ Douglas Palmer; Peter Barrett (2009). Evolution: The Story of Life. London: The Natural History Museum. ISBN 978-1-84533-339-3.
- ^ Dunne, Emma M.; Farnsworth, Alexander; Benson, Roger B. J.; Godoy, Pedro L.; Greene, Sarah E.; Valdes, Paul J.; Lunt, Daniel J.; Butler, Richard J. (9 January 2023). "Climatic controls on the ecological ascendancy of dinosaurs". Current Biology. 33 (1): 206–214. Bibcode:2023CBio...33E.206D. doi:10.1016/j.cub.2022.11.064. PMID 36528026. S2CID 254754419.
- ^ Scheyer et al. (2014): Early Triassic Marine Biotic Recovery: The Predators' Perspective. PLoS ONE https://doi.org/10.1371/journal.pone.0088987
- ^ Ruben, J. A. & Jones, T. D. (1 August 2000). "Selective Factors Associated with the Origin of Fur and Feathers". Integrative and Comparative Biology. 40 (4): 585–96. doi:10.1093/icb/40.4.585. Retrieved 24 November 2023.
- ^ Dai, Xu; Davies, Joshua H.F.L.; Yuan, Zhiwei; Brayard, Arnaud; Ovtcharova, Maria; Xu, Guanghui; Liu, Xiaokang; Smith, Christopher P.A.; Schweitzer, Carrie E.; Li, Mingtao; Perrot, Morgann G.; Jiang, Shouyi; Miao, Luyi; Cao, Yiran; Yan, Jia; Bai, Ruoyu; Wang, Fengyu; Guo, Wei; Song, Huyue; Tian, Li; Dal Corso, Jacopo; Liu, Yuting; Chu, Daoliang; Song, Haijun (2023). "A Mesozoic fossil lagerstätte from 250.8 million years ago shows a modern-type marine ecosystem". Science. 379 (6632): 567–572. Bibcode:2023Sci...379..567D. doi:10.1126/science.adf1622. PMID 36758082. S2CID 256697946.
- ^ Brayard, Arnaud; Krumenacker, L. J.; Botting, Joseph P.; Jenks, James F.; Bylund, Kevin G.; Fara, Emmanuel; Vennin, Emmanuelle; Olivier, Nicolas; Goudemand, Nicolas; Saucède, Thomas; Charbonnier, Sylvain; Romano, Carlo; Doguzhaeva, Larisa; Thuy, Ben; Hautmann, Michael; Stephen, Daniel A.; Thomazo, Christophe; Escarguel, Gilles (2017). "Unexpected Early Triassic marine ecosystem and the rise of the Modern evolutionary fauna". Science Advances. 3 (2): e1602159. Bibcode:2017SciA....3E2159B. doi:10.1126/sciadv.1602159. PMC 5310825. PMID 28246643.
- ^ Nomade et al., 2007 Palaeogeography, Palaeoclimatology, Palaeoecology 244, 326–44.
- ^ Marzoli et al., 1999, Science 284. Extensive 200-million-year-old continental flood basalts of the Central Atlantic Magmatic Province, pp. 618–620.
- ^ Hodych & Dunning, 1992.
- ^ "No Significant Nonmarine Carnian-Norian (Late Triassic) Extinction Event: Evidence From Petrified Forest National Park". gsa.confex.com. Archived from the original on 2003-11-06. Retrieved 2003-12-12.
References edit
- Emiliani, Cesare. (1992). Planet Earth: Cosmology, Geology, & the Evolution of Life & the Environment. Cambridge University Press. (Paperback Edition ISBN 0-521-40949-7)
- Ogg, Jim; June, 2004, Overview of Global Boundary Stratotype Sections and Points (GSSP's) Stratigraphy.org, Accessed April 30, 2006
- Stanley, Steven M. Earth System History. New York: W.H. Freeman and Company, 1999. ISBN 0-7167-2882-6
- Sues, Hans-Dieter & Fraser, Nicholas C. Triassic Life on Land: The Great Transition New York: Columbia University Press, 2010. Series: Critical Moments and Perspectives in Earth History and Paleobiology. ISBN 978-0-231-13522-1
- van Andel, Tjeerd, (1985) 1994, New Views on an Old Planet: A History of Global Change, Cambridge University Press
External links edit
- Overall introduction
- 'The Triassic world'
- Douglas Henderson's illustrations of Triassic animals[permanent dead link]
- Paleofiles page on the Triassic extinctions
- Examples of Triassic Fossils
- Triassic (chronostratigraphy scale)