


Summary
Color (American English) or colour (Commonwealth English) is the visual perception based on the electromagnetic spectrum. Though color is not an inherent property of matter, color perception is related to an object's light absorption, reflection, emission spectra and interference. For most humans, colors are perceived in the visible light spectrum with three types of cone cells (trichromacy). Other animals may have a different number of cone cell types or have eyes sensitive to different wavelength, such as bees that can distinguish ultraviolet, and thus have a different color sensitivity range. Animal perception of color originates from different light wavelength or spectral sensitivity in cone cell types, which is then processed by the brain.

Colors have perceived properties such as hue, colorfulness (saturation) and luminance. Colors can also be additively mixed (commonly used for actual light) or subtractively mixed (commonly used for materials). If the colors are mixed in the right proportions, because of metamerism, they may look the same as a single-wavelength light. For convenience, colors can be organized in a color space, which when being abstracted as a mathematical color model can assign each region of color with a corresponding set of numbers. As such, color spaces are an essential tool for color reproduction in print, photography, computer monitors and television. The most well-known color models are RGB, CMYK, YUV, HSL and HSV.
Because the perception of color is an important aspect of human life, different colors have been associated with emotions, activity, and nationality. Names of color regions in different cultures can have different, sometimes overlapping areas. In visual arts, color theory is used to govern the use of colors in an aesthetically pleasing and harmonious way. The theory of color includes the color complements; color balance; and classification of primary colors (traditionally red, yellow, blue), secondary colors (traditionally orange, green, purple) and tertiary colors. The study of colors in general is called color science.
Physical properties edit
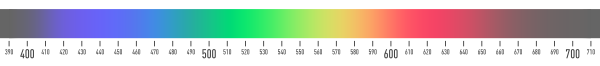
Electromagnetic radiation is characterized by its wavelength (or frequency) and its intensity. When the wavelength is within the visible spectrum (the range of wavelengths humans can perceive, approximately from 390 nm to 700 nm), it is known as "visible light".[1]
Most light sources emit light at many different wavelengths; a source's spectrum is a distribution giving its intensity at each wavelength. Although the spectrum of light arriving at the eye from a given direction determines the color sensation in that direction, there are many more possible spectral combinations than color sensations. In fact, one may formally define a color as a class of spectra that give rise to the same color sensation, although such classes would vary widely among different species, and to a lesser extent among individuals within the same species. In each such class, the members are called metamers of the color in question. This effect can be visualized by comparing the light sources' spectral power distributions and the resulting colors.
Spectral colors edit
The familiar colors of the rainbow in the spectrum—named using the Latin word for appearance or apparition by Isaac Newton in 1671—include all those colors that can be produced by visible light of a single wavelength only, the pure spectral or monochromatic colors. The spectrum above shows approximate wavelengths (in nm) for spectral colors in the visible range. Spectral colors have 100% purity, and are fully saturated. A complex mixture of spectral colors can be used to describe any color, which is the definition of a light power spectrum.
The spectral colors form a continuous spectrum, and how it is divided into distinct colors linguistically is a matter of culture and historical contingency.[2] Despite the ubiquitous ROYGBIV mnemonic used to remember the spectral colors in English, the inclusion or exclusion of colors is contentious, with disagreement often focused on indigo[broken anchor] and cyan.[3] Even if the subset of color terms is agreed, their wavelength ranges and borders between them may not be.
The intensity of a spectral color, relative to the context in which it is viewed, may alter its perception considerably according to the Bezold–Brücke shift; for example, a low-intensity orange-yellow is brown, and a low-intensity yellow-green is olive green. In color models capable of representing spectral colors,[4] such as CIELUV, a spectral color has the maximal saturation. In Helmholtz coordinates, this is described as 100% purity.
Color of objects edit
The physical color of an object depends on how it absorbs and scatters light. Most objects scatter light to some degree and do not reflect or transmit light specularly like glasses or mirrors. A transparent object allows almost all light to transmit or pass through, thus transparent objects are perceived as colorless. Conversely, an opaque object does not allow light to transmit through and instead absorbing or reflecting the light it receives. Like transparent objects, translucent objects allow light to transmit through, but translucent objects are seen colored because they scatter or absorb certain wavelengths of light via internal scatterance. The absorbed light is often dissipated as heat.[5]
Color vision edit
Development of theories of color vision edit

Although Aristotle and other ancient scientists had already written on the nature of light and color vision, it was not until Newton that light was identified as the source of the color sensation. In 1810, Goethe published his comprehensive Theory of Colors in which he provided a rational description of color experience, which 'tells us how it originates, not what it is'. (Schopenhauer)
In 1801 Thomas Young proposed his trichromatic theory, based on the observation that any color could be matched with a combination of three lights. This theory was later refined by James Clerk Maxwell and Hermann von Helmholtz. As Helmholtz puts it, "the principles of Newton's law of mixture were experimentally confirmed by Maxwell in 1856. Young's theory of color sensations, like so much else that this marvelous investigator achieved in advance of his time, remained unnoticed until Maxwell directed attention to it."[6]
At the same time as Helmholtz, Ewald Hering developed the opponent process theory of color, noting that color blindness and afterimages typically come in opponent pairs (red-green, blue-orange, yellow-violet, and black-white). Ultimately these two theories were synthesized in 1957 by Hurvich and Jameson, who showed that retinal processing corresponds to the trichromatic theory, while processing at the level of the lateral geniculate nucleus corresponds to the opponent theory.[7]
In 1931, an international group of experts known as the Commission internationale de l'éclairage (CIE) developed a mathematical color model, which mapped out the space of observable colors and assigned a set of three numbers to each.
Color in the eye edit

The ability of the human eye to distinguish colors is based upon the varying sensitivity of different cells in the retina to light of different wavelengths. Humans are trichromatic—the retina contains three types of color receptor cells, or cones. One type, relatively distinct from the other two, is most responsive to light that is perceived as blue or blue-violet, with wavelengths around 450 nm; cones of this type are sometimes called short-wavelength cones or S cones (or misleadingly, blue cones). The other two types are closely related genetically and chemically: middle-wavelength cones, M cones, or green cones are most sensitive to light perceived as green, with wavelengths around 540 nm, while the long-wavelength cones, L cones, or red cones, are most sensitive to light that is perceived as greenish yellow, with wavelengths around 570 nm.
Light, no matter how complex its composition of wavelengths, is reduced to three color components by the eye. Each cone type adheres to the principle of univariance, which is that each cone's output is determined by the amount of light that falls on it over all wavelengths. For each location in the visual field, the three types of cones yield three signals based on the extent to which each is stimulated. These amounts of stimulation are sometimes called tristimulus values.[8]
The response curve as a function of wavelength varies for each type of cone. Because the curves overlap, some tristimulus values do not occur for any incoming light combination. For example, it is not possible to stimulate only the mid-wavelength (so-called "green") cones; the other cones will inevitably be stimulated to some degree at the same time. The set of all possible tristimulus values determines the human color space. It has been estimated that humans can distinguish roughly 10 million different colors.[9]
The other type of light-sensitive cell in the eye, the rod, has a different response curve. In normal situations, when light is bright enough to strongly stimulate the cones, rods play virtually no role in vision at all.[10] On the other hand, in dim light, the cones are understimulated leaving only the signal from the rods, resulting in a colorless response. (Furthermore, the rods are barely sensitive to light in the "red" range.) In certain conditions of intermediate illumination, the rod response and a weak cone response can together result in color discriminations not accounted for by cone responses alone. These effects, combined, are summarized also in the Kruithof curve, which describes the change of color perception and pleasingness of light as a function of temperature and intensity.
Color in the brain edit
While the mechanisms of color vision at the level of the retina are well-described in terms of tristimulus values, color processing after that point is organized differently. A dominant theory of color vision proposes that color information is transmitted out of the eye by three opponent processes, or opponent channels, each constructed from the raw output of the cones: a red–green channel, a blue–yellow channel, and a black–white "luminance" channel. This theory has been supported by neurobiology, and accounts for the structure of our subjective color experience. Specifically, it explains why humans cannot perceive a "reddish green" or "yellowish blue", and it predicts the color wheel: it is the collection of colors for which at least one of the two color channels measures a value at one of its extremes.
The exact nature of color perception beyond the processing already described, and indeed the status of color as a feature of the perceived world or rather as a feature of our perception of the world—a type of qualia—is a matter of complex and continuing philosophical dispute.[citation needed]

From the V1 blobs, color information is sent to cells in the second visual area, V2. The cells in V2 that are most strongly color tuned are clustered in the "thin stripes" that, like the blobs in V1, stain for the enzyme cytochrome oxidase (separating the thin stripes are interstripes and thick stripes, which seem to be concerned with other visual information like motion and high-resolution form). Neurons in V2 then synapse onto cells in the extended V4. This area includes not only V4, but two other areas in the posterior inferior temporal cortex, anterior to area V3, the dorsal posterior inferior temporal cortex, and posterior TEO.[11][12] Area V4 was initially suggested by Semir Zeki to be exclusively dedicated to color,[13] and he later showed that V4 can be subdivided into subregions with very high concentrations of color cells separated from each other by zones with lower concentration of such cells though even the latter cells respond better to some wavelengths than to others,[14] a finding confirmed by subsequent studies.[11][15][16] The presence in V4 of orientation-selective cells led to the view that V4 is involved in processing both color and form associated with color[17] but it is worth noting that the orientation selective cells within V4 are more broadly tuned than their counterparts in V1, V2 and V3.[14] Color processing in the extended V4 occurs in millimeter-sized color modules called globs.[11][12] This is the part of the brain in which color is first processed into the full range of hues found in color space.[18][11][12]
Nonstandard color perception edit
Color vision deficiency edit
A color vision deficiency causes an individual to perceive a smaller gamut of colors than the standard observer with normal color vision. The effect can be mild, having lower "color resolution" (i.e. anomalous trichromacy), moderate, lacking an entire dimension or channel of color (e.g. dichromacy), or complete, lacking all color perception (i.e. monochromacy). Most forms of color blindness derive from one or more of the three classes of cone cells either being missing, having a shifted spectral sensitivity or having lower responsiveness to incoming light. In addition, cerebral achromatopsia is caused by neural anomalies in those parts of the brain where visual processing takes place.
Some colors that appear distinct to an individual with normal color vision will appear metameric to the color blind. The most common form of color blindness is congenital red–green color blindness, affecting ~8% of males. Individuals with the strongest form of this condition (dichromacy) will experience blue and purple, green and yellow, teal and gray as colors of confusion, i.e. metamers.[19]
Tetrachromacy edit
Outside of humans, which are mostly trichromatic (having three types of cones), most mammals are dichromatic, possessing only two cones. However, outside of mammals, most vertebrate are tetrachromatic, having four types of cones, and includes most, birds,[20][21][22] reptiles, amphibians and bony fish.[23][24] An extra dimension of color vision means these vertebrates can see two distinct colors that a normal human would view as metamers. Some invertebrates, such as the mantis shrimp, have an even higher number of cones (12) that could lead to a richer color gamut than even imaginable by humans.
The existence of human tetrachromats is a contentious notion. As many as half of all human females have 4 distinct cone classes, which could enable tetrachromacy.[25] However, a distinction must be made between retinal (or weak) tetrachromats, which express four cone classes in the retina, and functional (or strong) tetrachromats, which are able to make the enhanced color discriminations expected of tetrachromats. In fact, there is only one peer-reviewed report of a functional tetrachromat.[26] It is estimated that while the average person is able to see one million colors, someone with functional tetrachromacy could see a hundred million colors.[27]
Synesthesia edit
In certain forms of synesthesia, perceiving letters and numbers (grapheme–color synesthesia) or hearing sounds (chromesthesia) will evoke a perception of color. Behavioral and functional neuroimaging experiments have demonstrated that these color experiences lead to changes in behavioral tasks and lead to increased activation of brain regions involved in color perception, thus demonstrating their reality, and similarity to real color percepts, albeit evoked through a non-standard route. Synesthesia can occur genetically, with 4% of the population having variants associated with the condition. Synesthesia has also been known to occur with brain damage, drugs, and sensory deprivation.[28]
The philosopher Pythagoras experienced synesthesia and provided one of the first written accounts of the condition in approximately 550 BCE. He created mathematical equations for musical notes that could form part of a scale, such as an octave.[29]
Afterimages edit
After exposure to strong light in their sensitivity range, photoreceptors of a given type become desensitized.[30][31] For a few seconds after the light ceases, they will continue to signal less strongly than they otherwise would. Colors observed during that period will appear to lack the color component detected by the desensitized photoreceptors. This effect is responsible for the phenomenon of afterimages, in which the eye may continue to see a bright figure after looking away from it, but in a complementary color. Afterimage effects have also been used by artists, including Vincent van Gogh.
Color constancy edit
When an artist uses a limited color palette, the human visual system tends to compensate by seeing any gray or neutral color as the color which is missing from the color wheel. For example, in a limited palette consisting of red, yellow, black, and white, a mixture of yellow and black will appear as a variety of green, a mixture of red and black will appear as a variety of purple, and pure gray will appear bluish.[32]
The trichromatic theory is strictly true when the visual system is in a fixed state of adaptation.[33] In reality, the visual system is constantly adapting to changes in the environment and compares the various colors in a scene to reduce the effects of the illumination. If a scene is illuminated with one light, and then with another, as long as the difference between the light sources stays within a reasonable range, the colors in the scene appear relatively constant to us. This was studied by Edwin H. Land in the 1970s and led to his retinex theory of color constancy.[34][35]
Both phenomena are readily explained and mathematically modeled with modern theories of chromatic adaptation and color appearance (e.g. CIECAM02, iCAM).[36] There is no need to dismiss the trichromatic theory of vision, but rather it can be enhanced with an understanding of how the visual system adapts to changes in the viewing environment.
Reproduction edit

Color reproduction is the science of creating colors for the human eye that faithfully represent the desired color. It focuses on how to construct a spectrum of wavelengths that will best evoke a certain color in an observer. Most colors are not spectral colors, meaning they are mixtures of various wavelengths of light. However, these non-spectral colors are often described by their dominant wavelength, which identifies the single wavelength of light that produces a sensation most similar to the non-spectral color. Dominant wavelength is roughly akin to hue.
There are many color perceptions that by definition cannot be pure spectral colors due to desaturation or because they are purples (mixtures of red and violet light, from opposite ends of the spectrum). Some examples of necessarily non-spectral colors are the achromatic colors (black, gray, and white) and colors such as pink, tan, and magenta.
Two different light spectra that have the same effect on the three color receptors in the human eye will be perceived as the same color. They are metamers of that color. This is exemplified by the white light emitted by fluorescent lamps, which typically has a spectrum of a few narrow bands, while daylight has a continuous spectrum. The human eye cannot tell the difference between such light spectra just by looking into the light source, although the color rendering index of each light source may affect the color of objects illuminated by these metameric light sources.
Similarly, most human color perceptions can be generated by a mixture of three colors called primaries. This is used to reproduce color scenes in photography, printing, television, and other media. There are a number of methods or color spaces for specifying a color in terms of three particular primary colors. Each method has its advantages and disadvantages depending on the particular application.
No mixture of colors, however, can produce a response truly identical to that of a spectral color, although one can get close, especially for the longer wavelengths, where the CIE 1931 color space chromaticity diagram has a nearly straight edge. For example, mixing green light (530 nm) and blue light (460 nm) produces cyan light that is slightly desaturated, because response of the red color receptor would be greater to the green and blue light in the mixture than it would be to a pure cyan light at 485 nm that has the same intensity as the mixture of blue and green.
Because of this, and because the primaries in color printing systems generally are not pure themselves, the colors reproduced are never perfectly saturated spectral colors, and so spectral colors cannot be matched exactly. However, natural scenes rarely contain fully saturated colors, thus such scenes can usually be approximated well by these systems. The range of colors that can be reproduced with a given color reproduction system is called the gamut. The CIE chromaticity diagram can be used to describe the gamut.
Another problem with color reproduction systems is connected with the initial measurement of color, or colorimetry. The characteristics of the color sensors in measurement devices (e.g. cameras, scanners) are often very far from the characteristics of the receptors in the human eye.
A color reproduction system "tuned" to a human with normal color vision may give very inaccurate results for other observers, according to color vision deviations to the standard observer.
The different color response of different devices can be problematic if not properly managed. For color information stored and transferred in digital form, color management techniques, such as those based on ICC profiles, can help to avoid distortions of the reproduced colors. Color management does not circumvent the gamut limitations of particular output devices, but can assist in finding good mapping of input colors into the gamut that can be reproduced.
Additive coloring edit

Additive color is light created by mixing together light of two or more different colors.[37][38] Red, green, and blue are the additive primary colors normally used in additive color systems such as projectors, televisions, and computer terminals.
Subtractive coloring edit


Subtractive coloring uses dyes, inks, pigments, or filters to absorb some wavelengths of light and not others.[39] The color that a surface displays comes from the parts of the visible spectrum that are not absorbed and therefore remain visible. Without pigments or dye, fabric fibers, paint base and paper are usually made of particles that scatter white light (all colors) well in all directions. When a pigment or ink is added, wavelengths are absorbed or "subtracted" from white light, so light of another color reaches the eye.
If the light is not a pure white source (the case of nearly all forms of artificial lighting), the resulting spectrum will appear a slightly different color. Red paint, viewed under blue light, may appear black. Red paint is red because it scatters only the red components of the spectrum. If red paint is illuminated by blue light, it will be absorbed by the red paint, creating the appearance of a black object.
The subtractive model also predicts the color resulting from a mixture of paints, or similar medium such as fabric dye, whether applied in layers or mixed together prior to application. In the case of paint mixed before application, incident light interacts with many different pigment particles at various depths inside the paint layer before emerging.[40]
Structural color edit
Structural colors are colors caused by interference effects rather than by pigments. Color effects are produced when a material is scored with fine parallel lines, formed of one or more parallel thin layers, or otherwise composed of microstructures on the scale of the color's wavelength. If the microstructures are spaced randomly, light of shorter wavelengths will be scattered preferentially to produce Tyndall effect colors: the blue of the sky (Rayleigh scattering, caused by structures much smaller than the wavelength of light, in this case, air molecules), the luster of opals, and the blue of human irises. If the microstructures are aligned in arrays, for example, the array of pits in a CD, they behave as a diffraction grating: the grating reflects different wavelengths in different directions due to interference phenomena, separating mixed "white" light into light of different wavelengths. If the structure is one or more thin layers then it will reflect some wavelengths and transmit others, depending on the layers' thickness.
Structural color is studied in the field of thin-film optics. The most ordered or the most changeable structural colors are iridescent. Structural color is responsible for the blues and greens of the feathers of many birds (the blue jay, for example), as well as certain butterfly wings and beetle shells. Variations in the pattern's spacing often give rise to an iridescent effect, as seen in peacock feathers, soap bubbles, films of oil, and mother of pearl, because the reflected color depends upon the viewing angle. Numerous scientists have carried out research in butterfly wings and beetle shells, including Isaac Newton and Robert Hooke. Since 1942, electron micrography has been used, advancing the development of products that exploit structural color, such as "photonic" cosmetics.[41]
Cultural perspective edit
Colors, their meanings and associations can play a major role in works of art, including literature.[42]
Associations edit
Individual colors have a variety of cultural associations such as national colors (in general described in individual color articles and color symbolism). The field of color psychology attempts to identify the effects of color on human emotion and activity. Chromotherapy is a form of alternative medicine attributed to various Eastern traditions. Colors have different associations in different countries and cultures.[43]
Different colors have been demonstrated to have effects on cognition. For example, researchers at the University of Linz in Austria demonstrated that the color red significantly decreases cognitive functioning in men.[44] The combination of the colors red and yellow together can induce hunger, which has been capitalized on by a number of chain restaurants.[45]
Color plays a role in memory development too. A photograph that is in black and white is slightly less memorable than one in color.[46] Studies also show that wearing bright colors makes you more memorable to people you meet.
Terminology edit
Colors vary in several different ways, including hue (shades of red, orange, yellow, green, blue, and violet, etc), saturation, brightness. Some color words are derived from the name of an object of that color, such as "orange" or "salmon", while others are abstract, like "red".
In the 1969 study Basic Color Terms: Their Universality and Evolution, Brent Berlin and Paul Kay describe a pattern in naming "basic" colors (like "red" but not "red-orange" or "dark red" or "blood red", which are "shades" of red). All languages that have two "basic" color names distinguish dark/cool colors from bright/warm colors. The next colors to be distinguished are usually red and then yellow or green. All languages with six "basic" colors include black, white, red, green, blue, and yellow. The pattern holds up to a set of twelve: black, gray, white, pink, red, orange, yellow, green, blue, purple, brown, and azure (distinct from blue in Russian and Italian, but not English).
See also edit
- Chromophore
- Color analysis
- Color in Chinese culture
- Color mapping
- Complementary colors
- Impossible color
- International Color Consortium
- International Commission on Illumination
- Lists of colors (compact version)
- Neutral color
- Pearlescent coating including Metal effect pigments
- Pseudocolor
- Primary, secondary and tertiary colors
References edit
- ^ Bettini, Alessandro (2016). A Course in Classical Physics, Vol. 4 – Waves and Light. Springer. pp. 95, 103. ISBN 978-3-319-48329-0.
- ^ Berlin, B. and Kay, P., Basic Color Terms: Their Universality and Evolution, Berkeley: University of California Press, 1969.
- ^ Waldman, Gary (2002). Introduction to light: the physics of light, vision, and color. Mineola: Dover Publications. p. 193. ISBN 978-0486421186.
- ^ "Perceiving Color" (PDF). courses.washington.edu.
- ^ Berns, Roy S. (2019). Billmeyer and Saltzman's Principles of Color Technology. Fred W. Billmeyer, Max Saltzman (4th ed.). Hoboken, NJ: Wiley. pp. 5–9, 12. ISBN 978-1119366683. OCLC 1080250734.
- ^ Hermann von Helmholtz, Physiological Optics: The Sensations of Vision, 1866, as translated in Sources of Color Science, David L. MacAdam, ed., Cambridge: MIT Press, 1970.
- ^ Palmer, S.E. (1999). Vision Science: Photons to Phenomenology, Cambridge, MA: MIT Press. ISBN 0262161834.
- ^ Magazine, Nicola Jones, Knowable. "Color Is in the Eye, and Brain, of the Beholder". Scientific American. Retrieved 2022-11-08.
{{cite web}}: CS1 maint: multiple names: authors list (link) - ^ Judd, Deane B.; Wyszecki, Günter (1975). Color in Business, Science and Industry. Wiley Series in Pure and Applied Optics (3rd ed.). New York: Wiley-Interscience. p. 388. ISBN 978-0471452126.
- ^ "Under well-lit viewing conditions (photopic vision), cones ...are highly active and rods are inactive."Hirakawa, K.; Parks, T.W. (2005). "Chromatic Adaptation and White-Balance Problem" (PDF). IEEE International Conference on Image Processing 2005. IEEE ICIP. pp. iii-984. doi:10.1109/ICIP.2005.1530559. ISBN 0780391349. Archived from the original (PDF) on November 28, 2006.
- ^ a b c d Conway BR, Moeller S, Tsao DY (November 2007). "Specialized color modules in macaque extrastriate cortex". Neuron. 56 (3): 560–73. doi:10.1016/j.neuron.2007.10.008. PMC 8162777. PMID 17988638. S2CID 11724926. Archived from the original on 2022-10-10. Retrieved 2023-12-08.
- ^ a b c Conway BR, Tsao DY (October 2009). "Color-tuned neurons are spatially clustered according to color preference within alert macaque posterior inferior temporal cortex". Proceedings of the National Academy of Sciences of the United States of America. 106 (42): 18034–9. Bibcode:2009PNAS..10618034C. doi:10.1073/pnas.0810943106. PMC 2764907. PMID 19805195.
- ^ Zeki SM (April 1973). "Colour coding in rhesus monkey prestriate cortex". Brain Research. 53 (2): 422–7. doi:10.1016/0006-8993(73)90227-8. PMID 4196224.
- ^ a b Zeki S (March 1983). "The distribution of wavelength and orientation selective cells in different areas of monkey visual cortex". Proceedings of the Royal Society of London. Series B, Biological Sciences. 217 (1209): 449–70. Bibcode:1983RSPSB.217..449Z. doi:10.1098/rspb.1983.0020. PMID 6134287. S2CID 39700958.
- ^ Bushnell BN, Harding PJ, Kosai Y, Bair W, Pasupathy A (August 2011). "Equiluminance cells in visual cortical area v4". The Journal of Neuroscience. 31 (35): 12398–412. doi:10.1523/JNEUROSCI.1890-11.2011. PMC 3171995. PMID 21880901.
- ^ Tanigawa H, Lu HD, Roe AW (December 2010). "Functional organization for color and orientation in macaque V4". Nature Neuroscience. 13 (12): 1542–8. doi:10.1038/nn.2676. PMC 3005205. PMID 21076422.
- ^ Zeki S (June 2005). "The Ferrier Lecture 1995 behind the seen: the functional specialization of the brain in space and time". Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences. 360 (1458): 1145–83. doi:10.1098/rstb.2005.1666. PMC 1609195. PMID 16147515.
- ^ Zeki, S. (1980). "The representation of colours in the cerebral cortex". Nature. 284 (5755): 412–418. Bibcode:1980Natur.284..412Z. doi:10.1038/284412a0. ISSN 1476-4687. PMID 6767195. S2CID 4310049.
- ^ Flück, Daniel (19 January 2009). "Colorblind colors of confusion". Colblindor. Retrieved 14 November 2022.
- ^ Bennett, Andrew T. D.; Cuthill, Innes C.; Partridge, Julian C.; Maier, Erhard J. (1996). "Ultraviolet vision and mate choice in zebra finches". Nature. 380 (6573): 433–435. Bibcode:1996Natur.380..433B. doi:10.1038/380433a0. S2CID 4347875.
- ^ Bennett, Andrew T. D.; Théry, Marc (2007). "Avian Color Vision and Coloration: Multidisciplinary Evolutionary Biology" (PDF). The American Naturalist. 169 (S1): S1–S6. doi:10.1086/510163. ISSN 0003-0147. JSTOR 510163. S2CID 2484928.
- ^ Cuthill, Innes C.; Partridge, Julian C.; Bennett, Andrew T. D.; Church, Stuart C.; Hart, Nathan S.; Hunt, Sarah (2000). J. B. Slater, Peter; Rosenblatt, Jay S.; Snowdon, Charles T.; Roper, Timothy J. (eds.). Ultraviolet Vision in Birds. Advances in the Study of Behavior. Vol. 29. Academic Press. p. 159. doi:10.1016/S0065-3454(08)60105-9. ISBN 978-0-12-004529-7.
- ^ Bowmaker, James K. (September 2008). "Evolution of vertebrate visual pigments". Vision Research. 48 (20): 2022–2041. doi:10.1016/j.visres.2008.03.025. PMID 18590925. S2CID 52808112.
- ^ Vorobyev, M. (November 1998). "Tetrachromacy, oil droplets and bird plumage colours". Journal of Comparative Physiology A. 183 (5): 621–33. doi:10.1007/s003590050286. PMID 9839454. S2CID 372159.
- ^ Jameson, K.A.; Highnote, S.M.; Wasserman, L.M. (2001). "Richer color experience in observers with multiple photopigment opsin genes" (PDF). Psychonomic Bulletin and Review. 8 (2): 244–261 [256]. doi:10.3758/BF03196159. PMID 11495112. S2CID 2389566. Archived (PDF) from the original on 2013-10-04.
- ^ Jordan, G.; Deeb, S.S.; Bosten, J.M.; Mollon, J.D. (20 July 2010). "The dimensionality of color vision in carriers of anomalous trichromacy". Journal of Vision. 10 (8): 12. doi:10.1167/10.8.12. PMID 20884587.
- ^ Kershner, Kate (26 July 2016). "Lucky Tetrachromats See World With Up to 100 Million Colors". Retrieved 9 February 2022.
- ^ Brang, David (22 November 2011). "Survival of the Synesthesia Gene: Why Do People Hear Colors and Taste Words?". PLOS Biology. 9 (11): e1001205. doi:10.1371/journal.pbio.1001205. PMC 3222625. PMID 22131906.
- ^ "A Brief History of Synesthesia in the Arts". Retrieved 9 February 2022.
- ^ Gersztenkorn, D; Lee, AG (Jul 2, 2014). "Palinopsia revamped: A systematic review of the literature". Survey of Ophthalmology. 60 (1): 1–35. doi:10.1016/j.survophthal.2014.06.003. PMID 25113609.
- ^ Bender, MB; Feldman, M; Sobin, AJ (Jun 1968). "Palinopsia". Brain: A Journal of Neurology. 91 (2): 321–38. doi:10.1093/brain/91.2.321. PMID 5721933.
- ^ Depauw, Robert C. "United States Patent". Archived from the original on 6 January 2012. Retrieved 20 March 2011.
- ^ Walters, H. V. (1942). "Some Experiments on the Trichromatic Theory of Vision". Proceedings of the Royal Society of London. Series B, Biological Sciences. 131 (862): 27–50. Bibcode:1942RSPSB.131...27W. doi:10.1098/rspb.1942.0016. ISSN 0080-4649. JSTOR 82365. S2CID 120320368.
- ^ "Edwin H. Land | Optica". www.optica.org. Retrieved 2023-12-08.
- ^ Campbell, F. W. (1994). "Edwin Herbert Land. 7 May 1909-1 March 1991". Biographical Memoirs of Fellows of the Royal Society. 40: 197–219. doi:10.1098/rsbm.1994.0035. ISSN 0080-4606. JSTOR 770305. S2CID 72500555.
- ^ M.D. Fairchild, Color Appearance Models Archived May 5, 2011, at the Wayback Machine, 2nd Ed., Wiley, Chichester (2005).
- ^ MacEvoy, Bruce. "handprint : colormaking attributes". www.handprint.com. Retrieved 26 February 2019.
- ^ David Briggs (2007). "The Dimensions of Color". Archived from the original on 2015-09-28. Retrieved 2011-11-23.
- ^ "Molecular Expressions Microscopy Primer: Physics of Light and Color – Introduction to the Primary Colors". micro.magnet.fsu.edu. Retrieved 2023-12-08.
- ^ Williamson, Samuel J; Cummins, Herman Z (1983). Light and Color in Nature and Art. New York: John Wiley & Sons, Inc. pp. 28–30. ISBN 0-471-08374-7.
Thus subtractive color mixing laws that successfully describe how light is altered by nonspectral filters also describes how light is altered by pigments.
- ^ "Economic and Social Research Council: Science in the Dock, Art in the Stocks". Archived from the original on November 2, 2007. Retrieved 2007-10-07.
- ^ Westfahl, Gary (2005). The Greenwood Encyclopedia of Science Fiction and Fantasy: Themes, Works, and Wonders. Greenwood Publishing Group. pp. 142–143. ISBN 978-0313329517.
- ^ "Chart: Color Meanings by Culture". Archived from the original on 2010-10-12. Retrieved 2010-06-29.
- ^ Dzulkifli, Mariam; Mustafar, Muhammad (2013). "The Influence of Colour on Memory Performance: A Review". The Malaysian Journal of Medical Sciences. 20 (2): 3–9. doi:10.1016/j.chb.2010.06.010. S2CID 17764339.
- ^ "There's a sneaky reason why you always see red and yellow on fast food logos". Business Insider. Retrieved 2022-02-09.
- ^ Gnambs, Timo; Appel, Markus; Batinic, Bernad (2010). "Color red in web-based knowledge testing". Computers in Human Behavior. 26 (6): 1625–1631. doi:10.1016/j.chb.2010.06.010. S2CID 17764339.
External links edit
- Color at the Encyclopædia Britannica
- Maund, Barry. "Color". In Zalta, Edward N. (ed.). Stanford Encyclopedia of Philosophy.
- "Color". Internet Encyclopedia of Philosophy.